Carcinosomatidae

| Règne | Animalia |
|---|---|
| Embranchement | Arthropoda |
| Sous-embr. | Chelicerata |
| Ordre | † Eurypterida |
| Sous-ordre | † Eurypterina |
| Infra-ordre | † Diploperculata |
| Super-famille | † Carcinosomatoidea |
Størmer, 1934
Genres de rang inférieur
- † Carcinosoma Claypole, 1890
- † Eocarcinosoma
- † Eusarcana Strand, 1942
- † Rhinocarcinosoma
Carcinosomatidae (nom dérivé du genre type Carcinosoma, dont le nom signifie « corps de crabe ») est une famille d'Euryptéridés, un groupe éteint d'arthropodes aquatiques. Ils appartiennent à la superfamille Carcinosomatoidea, également nommée d'après Carcinosoma. Des fossiles de Carcinosomatides ont été découverts en Amérique du Nord, en Europe et en Asie, la famille ayant probablement colonisé le monde entier sur une période s'étendant de l'Ordovicien supérieur au Dévonien inférieur. Ils font partie des Euryptérides les plus "marins", les spécimens fossiles ayant presque tous été découverts dans des environnements marins.
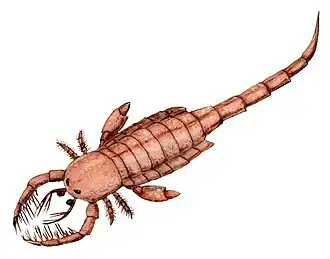
Les Carcinosomatides varient considérablement en taille, depuis des espèces mesurant seulement quelques centimètres de long jusqu'à certains des plus grands arthropodes connus. La plus grande espèce de Carcinosomatidé, Carcinosoma punctatum, mesure au moins 2,2 mètres (7,21784778 pi) et rivalise en taille avec le plus grand Euryptéride, Jaekelopterus. Morphologiquement, les Carcinosomatides sont très différents des autres Euryptéridés. Ils sont caractérisés par des membres antérieurs puissants et épineux, un corps central large et arrondi et une queue fine et tubulaire se terminant par un telson (la division la plus postérieure du corps) généralement courbé. Grâce à ces adaptations, les Carcinosomatides sont assez semblables aux scorpions, et le groupe a peut-être contribué au nom commun des Euryptérides, les « scorpions de mer ». La famille contient quatre, voire cinq, genres : Carcinosoma, Eocarcinosoma, Eusarcana, Rhinocarcinosoma et peut-être le genre problématique Holmipterus.
Il est peu probable que les Carcinosomatides aient été de puissants et actifs nageurs, compte tenu de leur forme non profilée. Il est plus probable qu'ils aient été nectobenthiques (nageant près du fond), possiblement superprédateurs (compte tenu de leur taille) ou charognards, creusant pour se nourrir ou pour creuser des terriers où se cacher pour chasser à l'affût.
Description

Les genres et espèces d'Euryptérides Carcinosomatides peuvent considérablement différer en taille, bien que la plupart des espèces soient assez grandes[1],[2]. La plus grande espèce est Carcinosoma punctatum avec 2,2 mètres (7,21784778 pi), soit l'un des plus grands Euryptérides, certains spécimens suggérant même qu'il aurait pu atteindre une longueur de 2,5 mètres (8,20209975 pi), rivalisant avec Jaekelopterus, le plus grand des Euryptérides[1],[3]. La plus petite espèce de Carcinosomatidé est Eusarcana obesus, avec 4 centimètres (1,5 en) de longueur[1] [note 1].
Morphologiquement, les Carcinosomatides se distinguent clairement des autres Eurypterides. Ce sont des Euryptérides nageurs (appartenant au sous-ordre des Eurypterina ), dotés de grandes palettes natatoires, d'un ensemble de membres antérieurs puissants et épineux, d'un préabdomen large et arrondi (corps central) et d'un abdomen mince, tubulaire, qui se termine par un telson (la division la plus postérieure du corps) de morphologie variable, souvent courbé[2],[7],[8],[9]. Dans un certain sens, les Carcinosomatides ressemblent plutôt à des scorpions, et ont peut-être contribué à ce que le nom commun des Eurypteridés soit devenu « scorpions de mer »[2],[10].
Il y a une grande variété de morphologie au sein du groupe. La carapace est de forme triangulaire à subtriangulaire chez tous les membres du groupe, mais la forme exacte peut varier. Rhinocarcinosoma, possède une protubérance distincte en forme de pelle à l'avant de la carapace[11]. Le préabdomen est large chez toutes les espèces, mais cette largeur diffère d'une espèce à l'autre. L'espèce relativement la plus large, est Eusarcana obesus, chez laquelle le quatrième segment est aussi large que les huit premiers segments combinés sont longs[12]. La spinosité (nombre d'épines) et la taille des membres antérieurs varient également d'un genre à l'autre, les membres antérieurs d'Eusarcana paraisant par exemple plus puissants que ceux de Rhinocarcinosoma[9]. Le telson varie considérablement entre les genres : chez Rhinocarcinosoma, il apparaît robuste et aplati, légèrement courbé vers le haut[9] ; chez Eusarcana, il est cylindrique et façonné en une pointe de queue acérée, semblable à celle d'un scorpion ; chez Carcinosoma, il est aplati, et se termine par une structure élargie et segmentée absente chez d'autres Euryptérides[8].
Histoire de la recherche

La première espèce de Carcinosomatidé à avoir été décrite est Carcinosoma punctatum, décrite pour la première fois sous le nom de Pterygotus punctatus par John William Salter en 1859[13]. Le premier genre considéré plus tard comme un Carcinosomatidé à avoir été décrit est Eusarcus (et son espèce type E. scorpionis ), décrit par August R. Grote et William Henry Pitt en 1875 sur la base de fossiles récupérés dans la chaux de Buffalo de l'âge Pridoli dans l'État de New York. Le genre n'ayant pas encore été décrit en tant que tel, et la description des fossiles se limitant au contour et à la forme, Henry Woodward attribue alors E. scorpionis à Eurypterus au motif que l'espèce est similaire en forme à Eurypterus punctatus (Pterygotus punctatus ayant été reclassé comme une espèce d' Eurypterus ). À l'insu de Grote et Pitt, Eusarcus se voit attribuer le même nom qu'un genre d'Opilione Laniatore contemporain de la famille des Gonyleptidae, décrit en 1833 et constitue donc un homonyme. L'homonymie passe inaperçue jusqu'aux années 1930[2]. Le genre Eurysoma est également décrit à la fin du XIXe siècle, aux côtés de son espèce type, E. newlini, par Edward Waller Claypole en 1890. Lorsque Claypole découvre plus tard en 1890 que le nom est déjà utilisé pour un genre de coléoptères modernes, il remplace le nom Eurysoma par le nom Carcinosoma[14].
En 1912, John Mason Clarke et Rudolf Ruedemann déclarent que les différences entre Eusarcus et toutes les formes apparentées d'Euryptéridés sont si grandes qu'il est « tout à fait évident » qu'Eusarcus est distinct des autres Euryptéridés. Clarke et Ruedemann attribuent plusieurs nouvelles espèces à Eusarcus, y compris de nouvelles espèces qui seront plus tard considérées comme appartenant au genre Rhinocarcinosoma. Ils concluent qu'Eusarcus est suffisamment similaire à Carcinosoma pour que les deux genres soient considérés comme synonymes. Eusarcus ayant été décrit avant Carcinosoma, les lois taxonomiques de priorité dictent qu'Eusarcus sera le nom du taxon[2].
En 1934, Leif Størmer se rend finalement compte qu'Eusarcus est un nom déjà utilisé. Il le remplace par le nom de son synonyme le plus ancien non encore utilisé, Carcinosoma. Størmer décrit également en 1934 la famille Carcinosomatidae, initialement sous le nom de « Carcinosomidae », pour regrouper les quatre genres Carcinosoma, Mixopterus, Echinognathus et Megalograptus. En 1942, Embrik Strand propose un autre nom de remplacement pour Eusarcus, Eusarcana, bien que la question ait déjà été traitée par Størmer huit ans auparavant[2]. La famille est modifiée par Erik N. Kjellesvig-Waering dans le Treatise on Invertebrate Paleontology de Størmer de 1955 : le nom devient définitivement Carcinosomatidae ; les genres autres que Carcinosoma sont transférés à leurs propres familles (Mixopterus aux Mixopteridae ; Megalograptus et Echinognathus aux Megalograptidae). En 1962, Rhinocarcinosoma est séparé de Carcinosoma par Nestor Ivanovich Novozhilov, sa carapace étant différente de celle des autres Carcinosoma[9].

A l'occasion d'une revue des Carcinosomatides en 1964, Kenneth Edward Caster et Erik N. Kjellesvig-Waering reconnaissent qu'Eusarcus et Carcinosoma sont des genres distincts, considérant comme erronée la synonymisation de 1912. Le nom Eusarcus étant déjà utilisé, Caster et Kjellesvig-Waering, qui ignorent probablement l'existence de l'Eusarcana de Strand, inventent un nouveau nom nom, Paracarcinosoma, pour l'espèce précédemment appelée Eusarcus[2]. La même année, Caster et Kjellesvig-Waering décrivent le nouveau genre Eocarcinosoma pour prendre en compte les spécimens fossiles d'Eusarcus/Paracarcinosoma de l'Ordovicien[15]. Bien que la plupart de ces spécimens aient depuis été identifiés comme des pseudo-fossiles, le spécimen type d'Eocarcinosoma est un fossile authentique et le plus ancien spécimen de la famille[16],[17],[18]. L'aire de répartition géographique connue des Carcinosomatides a été considérablement étendue par la découverte de fossiles de Rhinocarcinosoma au Vietnam à la fin du 20e siècle, nommés R. dosonensis en 2002[9],[19].
En 2012, Jason A. Dunlop et James Lamsdell reconnaissent le nom Eusarcana, le premier apparu, comme le nom de remplacement valide d'Eusarcus[2]. Ils transfèrent l'espèce attribuée à Paracarcinosoma à ce genrer[2]. Ils désignent Paracarcinosoma comme synonyme junior[2]. Le nom Paracarcinosoma continue néanmoins à être fréquemment utilisé par des chercheurs. En 2015, une analyse phylogénétique réalisée par Lamsdell et ses collègues permet de d'identifier Holmipterus, un Carcinosomatide basal, genre d'Euryptéride problématique aux affinités incertaines[20]. Toutefois, la position de Holmipterus est loin d'être certaine dans l'arbre généalogique des Euryptéridés, en raison d'un matériel fossile incomplet et d'une combinaison apparente de traits de différentes familles[21]. Ses fossiles pourraient même représenter deux genres différents, regroupés par erreur[22].
Classification
Les Carcinosomatides sont classés dans la super-famille Carcinosomatoidea, au sein de l'infra-ordre Diploperculata[18]. Les Carcinosomatoidea comprennent également les familles Mixopteridae et Megalograptidae[20],[18]. Auparavant, de 1989 au début des années 2000, les Carcinosomatidae étaient regroupés avec la famille Hughmilleriidae dans la super-famille Hughmillerioidea, parce qu'ils partageaient des membres épineux et étaient considérés d'un type cohérent sauf en ce qui concernaient les palettes natatoires[22],[23]. Les Hughmilleriidae sont aujourd'hui considérés comme des membres basaux de la super-famille des Pterygotioidea[24].
La phylogénie interne des Carcinosomatoidea est mal résolue (peu claire)[10]. Le premier cladogramme ci-dessous est issu d'une étude de 2007 conduite par le chercheur sur les Euryptéridés, O. Erik Tetlie, étude elle même basée sur les résultats de diverses analyses phylogénétiques sur les Euryptéridés menées entre 2004 et 2007[10]. Le deuxième cladogramme est issu d'une étude de 2015 menée par James Lamsdell et ses collègues[20]. Les deux cladogrammes ont été simplifiés pour n'afficher que les Carcinosomatoidea. Tetlie (2007) classe les Carcinosomatidae comme un groupe paraphylétique, représentant les membres basaux des Carcinosomatoidea[10]. Lamsdell et al . (2015) positionnent les Carcinosomatides comme un groupe monophylétique[20].
Tetlie (2007)
| Carcinosomatoidea |
| ||||||||||||||||||||||||||||||||||||||||||
Lamsdell et al. (2015)
| Carcinosomatoidea |
| ||||||||||||||||||||||||||||||||||||||||||
Paléoécologie
.jpg)
Les Euryptérides Carcinosomatides comptent parmi les Euryptérides les plus marins[19]. Leurs fossiles proviennent de dépôts qui étaient autrefois des récifs, dans des environnements lagunaires, ou dans des eaux plus profondes[10],[11]. Cela contraste nettement avec leurs plus proches parents, les Mixoptérides, qu'on ne retrouve pas dans les milieux d'eaux profondes. La seule autre famille d'Euryptérides connue pour avoir occupé des milieux d'eaux profondes est celle des Ptérygotides, qui ont une distribution similaire à celle des Carcinosomatides, bien que plus réussie. En se basant sur la répartition des Ptérygotides, on peut inférer une répartition cosmopolite pour les Carcinosomatides. Ils constituent, avec les Ptérygotides, l'une des deux seules familles d'Euryptérides connues du continent austral Gondwana du Silurien au Dévonien[10]. Le seul genre de Carcinosomatide connu dans des dépôts non marins est Rhinocarcinosoma, découvert dans des dépôts fluviaux (rivières) et lacustres (lacs), bien que des fossiles du genre aient également été découverts dans des dépôts marins[19].
Leurs corps n'étant pas aussi profilés que ceux de nombreux autres Euryptérides nageurs, et compte tenu de la morphologie singulière de leurs telsons, il est probable que les Carcinosomatides n'étaient pas des nageurs très actifs, et qu'ils aient adopté un mode de vie plus nektobenthique (nage près du fond)[13]. Les caractères singuliers de Rhinocarcinosoma illustrent bien l'adaptation à ce mode de vie, en particulier la protubérance en forme de pelle à l'avant de sa carapace qui pourrait avoir été utilisée pour creuser ou fouiller la vase, et les palettes natatoires de taille réduite par rapport à celles des autres Carcinosomatides[19]. Compte tenu de leur taille, les Carcinosomatides étaient peut-être des superprédateurs ou des charognards, creusant pour se nourrir ou pour s'aménager des terriers pour chasser à l'affût. Ils se nourrissaient peut-être de vers, d'autres arthropodes, de brachiopodes et de poissons, utilisant leurs membres antérieurs pour pousser la nourriture dans leur bouche[19].
Liens externes
- Ressources relatives au vivant :
Notes et références
Notes
- ↑ Les plus grands spécimens fossiles de l'espèce Rhinocarcinosoma cicerops font aussi 4 centimètres de long, mais les fossiles connus représentent des formes juvéniles[1],[4]. Eocarcinosoma batrachophthalmus était aussi très petit, mais il aurait dépassé les 4 centimètres de long compte tenu du fait que sa tête faisait 2.05 centimètres (0.8 in) de long[5]. Il est aussi possible qu'Eusarcana obesus soit une forme juvénile de l'espèce Carcinosoma scorpioides[6], et dans ce cas Eocarcinosoma batrachophthalmus serait le plus petit Carcinosomatide[1].
Références
- 1 2 3 4 5 (en) Lamsdell et Braddy, « Cope's rule and Romer's theory: patterns of diversity and gigantism in eurypterids and Palaeozoic vertebrates », Biology Letters, vol. 6, no 2, , p. 265–269 (ISSN 1744-9561, PMID 19828493, PMCID 2865068, DOI 10.1098/rsbl.2009.0700, lire en ligne)
- 1 2 3 4 5 6 7 8 9 10 (en) Dunlop et Lamsdell, « Nomenclatural notes on the eurypterid family Carcinosomatidae », Zoosystematics and Evolution, vol. 88, no 1, , p. 19–24 (ISSN 1860-0743, DOI 10.1002/zoos.201200003, lire en ligne)
- ↑ (en) Kjellesvig-Waering, « The Silurian Eurypterida of the Welsh Borderland », Journal of Paleontology, vol. 35, no 4, , p. 789–835 (JSTOR 1301214)
- ↑ John M. Clarke et Rudolf Ruedemann, The Eurypterida of New York, University of California Libraries, (ISBN 978-1125460221, lire en ligne)
- ↑ (en) Kenneth Edward Caster et Erik N. Kjellesvig-Waering, « Upper Ordovician eurypterids of Ohio », Paleontological Research Institution, vol. 4, (lire en ligne)
- ↑ (en) James Lamsdell, « Selectivity in the evolution of Palaeozoic arthropod groups, with focus on mass extinctions and radiations: a phylogenetic approach », Submitted to the graduate degree program in Geology and the Graduate Faculty of the University of Kansas in partial fulfillment of the requirements for the degree of Doctor of Philosophy, (lire en ligne)
- ↑ Erik N. Kjellesvig-Waering, Lower Wenlock faunal and floral dynamics – Vattenfallet section, Gotland, Geological Survey of Sweden, , 121–136 p. (ISBN 9171581707, lire en ligne), « Eurypterids »
- 1 2 (en) Tetlie, « Eurypterida (Chelicerata) from the Welsh Borderlands, England », Geological Magazine, vol. 143, no 5, , p. 723–735 (ISSN 1469-5081, DOI 10.1017/S0016756806002536, Bibcode 2006GeoM..143..723T, S2CID 83835591)
- 1 2 3 4 5 (en) Braddy, Selden et Truong, « A New Carcinosomatid Eurypterid From The Upper Silurian Of Northern Vietnam », Palaeontology, vol. 45, no 5, , p. 897–915 (ISSN 1475-4983, DOI 10.1111/1475-4983.00267, hdl 1808/8358, S2CID 129450304, lire en ligne)
- 1 2 3 4 5 6 (en) Tetlie, « Distribution and dispersal history of Eurypterida (Chelicerata) », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 252, nos 3–4, , p. 557–574 (DOI 10.1016/j.palaeo.2007.05.011, lire en ligne [archive du ])
- 1 2 (en) Ciurca, « New occurrences of Silurian eurypterids (Carcinosomatidae) in Pennsylvania, Ohio and New York », The Paleontological Society Special Publications, vol. 6, , p. 57 (ISSN 2475-2622, DOI 10.1017/S2475262200006171)
- ↑ (en) Woodward, « On some New Species of Crustacea from the Upper Silurian Rocks of Lanarkshire &c.; and further observations on the Structure of Pterygotus », Quarterly Journal of the Geological Society, vol. 24, nos 1–2, , p. 289–296 (ISSN 0370-291X, DOI 10.1144/GSL.JGS.1868.024.01-02.36, S2CID 128874377)
- 1 2 (en) Gladwell, « The biota of Upper Silurian submarine channel deposits, Welsh Borderland », Theses, Leicester University Dept. Of Geology, (lire en ligne)
- ↑ Kjellesvig-Waering, « Some Previously Unknown Morphological Structures of Carcinosoma newlini (Claypole) », Journal of Paleontology, vol. 32, no 2, , p. 295–303 (JSTOR 1300736)
- ↑ Caster et Kjellesvig-Waering, « Upper Ordovician eurypterids of Ohio », Paleontological Research Institution, vol. 4, (lire en ligne)
- ↑ Tollerton, « Summary of a revision of New York State Ordovician eurypterids: implications for eurypterid palaeoecology, diversity and evolution », Earth and Environmental Science Transactions of the Royal Society of Edinburgh, vol. 94, no 3, , p. 235 (ISSN 0263-5933, DOI 10.1017/s0263593303000154, lire en ligne)
- ↑ (en) Simon J. Braddy, Victor P. Tollerton, Patrick R. Racheboeuf et Roger Schallreuter, 25. Eurypterids, Phyllocarids, and Ostracodes, Columbia University Press, (ISBN 978-0-231-50163-7, DOI 10.7312/webb12678-026, lire en ligne)
- 1 2 3 (en) J. A. Dunlop, D. Penney et D. Jekel, World Spider Catalog : A summary list of fossil spiders and their relatives, Musée d'histoire naturelle de Berne, (lire en ligne)
- 1 2 3 4 5 Thanh, Janvier, Truong et Braddy, « New vertebrate remains associated with Eurypterids from the Devonian Do Son Formation Vietnam », Journal of Geology, vol. 3–4, , p. 1–11 (lire en ligne)
- 1 2 3 4 Lamsdell, Briggs, Liu et Witzke, « The oldest described eurypterid: a giant Middle Ordovician (Darriwilian) megalograptid from the Winneshiek Lagerstätte of Iowa », BMC Evolutionary Biology, vol. 15, , p. 169 (PMID 26324341, PMCID 4556007, DOI 10.1186/s12862-015-0443-9)
- ↑ Erik N. Kjellesvig-Waering, Lower Wenlock faunal and floral dynamics – Vattenfallet section, Gotland, Geological Survey of Sweden, , 121–136 p. (ISBN 9171581707, lire en ligne), « Eurypterids »
- 1 2 (en) Tollerton, « Morphology, taxonomy, and classification of the order Eurypterida Burmeister, 1843 », Journal of Paleontology, vol. 63, no 5, , p. 642–657 (ISSN 0022-3360, DOI 10.1017/S0022336000041275, S2CID 46953627, lire en ligne)
- ↑ (en) Stott, Tetlie, Braddy et Nowlan, « A new eurypterid (Chelicerata) from the Upper Ordovician of Manitoulin Island, Ontario, Canada », Journal of Paleontology, vol. 79, no 6, , p. 1166–1174 (ISSN 0022-3360, DOI 10.1666/0022-3360(2005)079[1166:ANECFT]2.0.CO;2, S2CID 55014027, lire en ligne)
- ↑ (en) Tetlie et Briggs, « The origin of pterygotid eurypterids (Chelicerata: Eurypterida) », Palaeontology, vol. 52, no 5, , p. 1141–1148 (ISSN 1475-4983, DOI 10.1111/j.1475-4983.2009.00907.x)
 Portail de la paléontologie
Portail de la paléontologie